
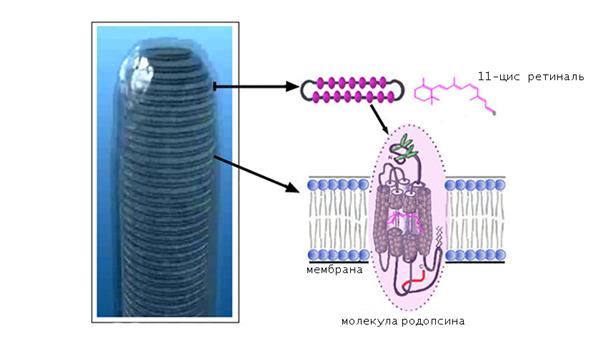
Родопсин — це поширений зоровий пігмент, що входить до складу паличковидних зорових рецепторів сітківки очі хребетних тварин. Ця речовина має дуже високу фоточутливість і є ключовим компонентом фоторецепції. Інша назва родопсину — зоровий пурпур.
Зараз до родопсинам відносять пігменти не тільки паличок, але і рабдомерных зорових рецепторів членистоногих.
Загальна характеристика пігменту
По хімічної природі родопсин — це мембранний білок тваринного походження, що містить у своїй структурі хромофорную групу. Саме вона зумовлює здатність пігменту поглинати кванти світла. Білок родопсин має молекулярну масу приблизно 40 кДА і містить 348 амінокислотних ланок.

Спектр світлопоглинання родопсину складається з трьох смуг:
- α (500 нм);
- β (350 нм);
- γ (280 нм).
Промені γ поглинаються ароматичними амінокислотами в складі поліпептидного ланцюга, а β і α — хромофорной групою.
Родопсин — це речовина, здатна розпадатися під дією світла, що запускає электротонический шлях передачі сигналу по нервовим волокнам. Дана властивість характерно і для інших пігментів фоторецепторів.
